企业QQ
官方微信

苏州蚂蚁淘生物科技有限公司成立于2017年,是一家专业提供实验室整体采购方案的专业公司,为广大采购和实验室解决了大量问题,几年来已经成为所有合作伙伴最优秀的合作伙伴,我们一直秉承信誉第一的原则,坚持不卖假货,哪怕利润损失也要为客户提供最好的产品
我们已经和国外500多个品牌建立了贸易往来,并在国际上享有相当知名度,现已经成为国外近100多个品牌中国指定代理,并成为其中国最优秀的代理商
亚基合成
图1.核糖核苷向吗啉代转化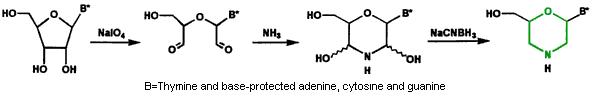
图2.吗啉亚基的保护和激活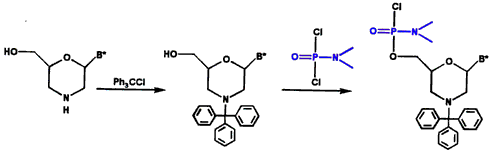
Oligo大会
图3. Oligo组装周期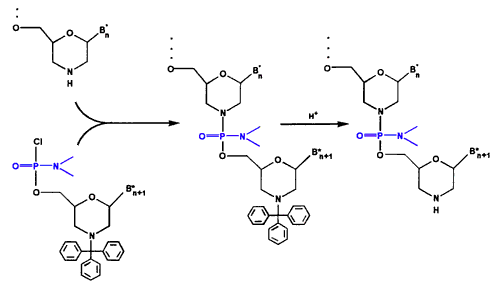
寡核苷酸加工
苏州蚂蚁淘生物科技有限公司SuZhouebiomall BIOLOGicalTechnologyCo.,Ltd.成立于2017年,经过数年的努力已迅速成为集科研和生产为一体的专业化生物工程公司。特别是ELISA研发,代测,技术服务这一产品已使苏州蚂蚁淘成为中国知名公司。公司产品涵盖ELISA研发、细胞培养,各种生化试剂、各种酶类、分子生物学试剂盒、培养基、自产实验室耗材、小型仪器等。此外、苏州蚂蚁淘还代理了美国药典标准品,中检所标准品,sigma,ScienCell,ATCC,wak0,omega,Worthington、MBI、Bioworld、Calendon等国外各类知名公司的产品。苏州蚂蚁淘将进一步加强人才经营、产品经营,不断提升企业的核心竟争力,实现具有高科技产业体系、知识化创业团队的国际化的公司,服务于生命科学领域,迎接世界经济一体化所带来的机遇与挑战。
Gene Tools选择最佳目标基因工具客户支持人员拥有工具和技术来选择可能产生良好敲除或剪接修饰的吗啉寡核苷酸序列。 Morpholino寡核苷酸靶向是Gene Tools提供的免费服务。将目标mRNA序列提交进行设计后,通常在下一个工作日通过电子邮件返回建议的Morpholino反义序列。介绍靶向Morpholino寡核苷酸比靶向基于DNA的反义寡核苷酸要简单得多。如果将吗啉成功地递送到细胞质中,通常会进行首次尝试。DNA和S-DNA寡核苷酸在首次尝试中关闭目标基因的可能性很小;在找到有效序列之前,通常尝试五到六个寡核苷酸。部分原因是吗啉寡核苷酸具有更高的结合亲和力而不是等同的基于DNA的反义寡核苷酸。这种较高的亲和力使Morpholinos能够侵入RNA二级结构,并大大提高了设计有效寡核苷酸的可能性。精确靶向单个基因也得到了简化,因为吗啉代寡核苷酸通过位阻作用而不是RNAse H介导的mRNA降解机制。我们将在本节中进一步描述亲和力和位阻效应。回答有关您的实验系统的几个问题,将有助于您设计针对目标靶点的特异且高效的吗啉序列。设计出色的Morpholino寡核苷酸并不需要所有这些问题的详尽答案,但是,您对目标的了解越多,成功的机会就越大。相关问题包括:mRNA或pre-mRNA的结构是什么?我想阻止剪接形式或剪接位点吗?我在起始密码子和上游是否有足够的序列来设计一个或多个翻译阻断型吗啉寡核苷酸?靶序列是否与不良基因靶具有同源性?我的学习有哪些实验条件(例如温度和时间限制)?除了上述定位问题之外,您应该牢记这两个对于总体实验设计很重要的问题:如何检测成功的基因敲低?我如何将寡核苷酸递送到细胞质中?一旦找到了可行的靶标区域并选择了有效的胞质递送方法和用于测定反义活性的方法,Morpholino寡核苷酸序列将决定成功。重要的是,寡核苷酸必须具有几乎没有自身互补性的碱基序列,以使其不二聚。否则,最佳的吗啉代寡核苷酸将长约25个碱基,并具有足够高的GC含量(例如40-60%),从而具有很高的目标亲和力。寡聚物不应具有四个或更多的连续G链段,以保持水溶性。在最典型的情况下,完整的mRNA序列是已知的,人们希望阻止其翻译。对于此应用,最佳靶标是25个碱基的序列,该序列位于5'帽到编码序列的前25个碱基的区域内,GC含量约为50%,几乎没有或没有二级结构。除了具有内部核糖体进入位点(IRES)的基因的异常情况外,靶向起始密码子而不是上游序列没有特别的优势。通常,人们可以在5'帽以内找到至少2或3个寡核苷酸靶,以起始具有大致相同性质的区域。与任何这些靶标互补的寡核苷酸应减少下游编码区的表达。在针对给定基因的任何寡核苷酸中,很可能存在“最佳”寡核苷酸,其可实现最大的敲除,有时达到无法检测到的基因表达水平。在许多情况下,仅尝试一个寡核苷酸就足以有效击倒。没有其他反义结构类型可以达到Morpholino寡核苷酸使用者成功完成首次尝试敲除的高比例。Morpholino寡核苷酸靶向的成功率很高,可能是由于它们对RNA的亲和力更高。与低亲和力的S-DNA相比,Morpholinos入侵RNA的二级结构要成功得多。反义领域的一个长期问题是难以预测反义寡核苷酸的有效靶标。具体而言,S-DNA对所选RNA中的许多潜在靶序列无活性。这可能是因为弱结合的S-DNA无法侵入RNA中相对稳定的二级结构。即使当RNA中合适的单链区域可用于配对时,低亲和力S-DNA常常也无法侵入相邻的双链区域。通常,对几套S-DNA寡核苷酸进行筛选,以找到能够敲除其靶基因的单个寡核苷酸。通常,Morpholino用户购买了一个寡核苷酸并敲低了一个基因;大约80%的时间,第一个Morpholino尝试执行此操作。Morpholinos的良好靶向可预测性通过图1a和1b中所示的实验证明。这些实验中靶向的mRNA具有相当稳定的二级结构,但吗啉代寡核苷酸仍能实现良好的敲低。图1a显示了包含与萤火虫荧光素酶的氨基酸编码序列连接的乙型肝炎病毒(HBV)mRNA前导序列的85个碱基的mRNA构建体。编号的线表示7种不同的吗啉代反义寡核苷酸的靶位点。图1b显示了在无细胞翻译测定中使用1纳摩尔靶标mRNA实现的翻译敲低。图1a中显示的每种Morpholino寡核苷酸均以1微摩尔浓度使用,其击倒功效在图1b中作为其沿mRNA位置的函数绘制,以及与平移起始位点相距3'的另外5个寡核苷酸的对应值。尤其值得注意的是,寡核苷酸3、4和5被认为有效侵入了HBV前导序列内相当稳定的二级结构。图1. RNA结构和吗啉反义活性应当指出,Morpholinos靶向AUG翻译起始位点3个以上碱基的低效力可能是在翻译起始位点完全组装后与核糖体复合物相关的解旋酶活性的结果。如我们在特异性部分中所述,针对翻译起始位点3个以上碱基的序列缺乏效力实际上实际上导致了Morpholinos相对于S-DNA,RNAi和嵌合寡聚物的特异性更高。图1b中的结果表明,通过遵循合理且可靠的靶向指南,可以选择具有较高有效可能性的吗啉代反义寡核苷酸。这些定位准则将在下面详细介绍。定位准则现在,我们将带您完成设计翻译阻断性Morpholino寡核苷酸的步骤。然后,我们将向您展示如何设计Morpholino寡核苷酸以阻断或改变mRNA剪接事件。当研究人员要求我们提供免费的oligo设计服务时,这些定位准则与Gene Tools客户支持人员所使用的准则相同。 阻断翻译:以下指导原则通常导致吗啉代寡核苷酸,其对阻断其靶mRNA的翻译非常有效。 1.在剪接后的mRNA中从5'帽到AUG翻译起始位点约25个碱基3'的区域中选择一个靶序列。如图2所示,Morpholinos(通过空间阻滞机制起作用)靶向AUG翻译起始位点3'左右约30个碱基,不会阻止翻译。图2.反义活性与mRNA中的靶位只要不存在禁止性的自我互补性或其他不良构想,包含翻译起始位点的25个碱基的区域就是一个简单且通常有效的目标。当基因工具设计了用于翻译阻断的Morpholino寡核苷酸,我们首先分析编码序列的前25个碱基(包括起始密码子),然后向上游滑动25个碱基的窗口,直到找到满足最佳常规要求的25个碱基Morpholino寡核苷酸(即GC含量为40-60%,无明显的自互补性或4个或更多连续G链的延伸)。以这种方式接近寡核苷酸靶向的优势在于,其他可行的靶标可能仍保留在5'UTR的上游。当研究人员希望用第二个或第三个不重叠的吗啉代寡核苷酸针对相同的靶mRNA验证其反义结果时,这一点就变得很重要。我们发现,许多发育生物学研究人员更喜欢这种“ 2至3寡核苷酸策略”来验证其反义结果。图3. mRNA的翻译阻断的可靶向区域的初始选择2.确保所选序列几乎没有或没有自身互补性。应该选择吗啉代序列,以使自身互补序列的稳定性最小。所选的吗啉代寡核苷酸应形成不超过16个连续的链内氢键,最好不超过16个。在某些情况下,如果所得的配对完全在G和C残基之间,则16个连续的碱基对可能不利于获得良好的反义结果。图4显示了寡核苷酸A,其几乎不能接受的16个连续碱基对的自互补性(2 + 3 + 3 + 3 + 3 + 3 + 2 = 16个连续氢键)和寡核苷酸B,不应选择,因为它含有自互补性的不可接受的18个连续碱基对(3 + 3 + 3 + 3 + 3 + 3 + 3 = 18个连续氢键)。图4.具有不同自我互补性的寡核苷酸注意,这些图显示了链内配对事件,并且寡聚B也将通过链间配对形成吗啉二聚体。3.如果一个25碱基的Morpholino寡核苷酸含有9个以上鸟嘌呤(> 36%G)或3个以上连续鸟嘌呤,则水溶性可能较差。4.对于大多数应用,建议使用25米(市面上最长的)。这是因为效率随着长度的增加而显着增加,并且因为长寡核苷酸最好确保接近目标RNA中的单链区域,这是寡核苷酸配对成核所必需的。图5证明了在无细胞翻译系统中,随着吗啉代寡核苷酸长度的增加,功效大大提高。图5.吗啉代寡核苷酸的长度与活性用含有尿嘧啶的吗啉代寡核苷酸进行了该长度对活性的研究。1998年,Gene Tools从尿嘧啶转变为胸腺嘧啶,从而增加了靶标结合亲和力,因此“新”含胸腺嘧啶的25-mer与“老”含尿嘧啶28-mer具有相同的功效。Morpholino寡核苷酸继续与胸腺嘧啶合成。从这项研究中可以看出,较短的吗啉(短于约20个亚基)仅显示适度的功效,而较长的吗啉(长达25-mer可用)则更有效。与反义领域的传统看法相反,增加吗啉的长度不会导致其特异性的重大损失。这在我们关于吗啉代寡核苷酸特异性的章节中有详细描述。与吗啉代寡核苷酸或其他空间封闭寡核苷酸相反,S-DNA寡核苷酸和其他依赖RNAse-H的寡核苷酸确实会随着长度的增加而显着降低特异性。阻止核处理:吗啉代寡核苷酸可以阻断核加工事件,特别是mRNA之前的剪接。事实证明,以下指南可有效设计针对剪接位点的寡核苷酸。与翻译阻断相比,用吗啉寡核苷酸阻断核加工具有一些明显的优势。首先,您通常可以通过遵循一些通用的寡核苷酸设计指南来创建特定的所需效果,例如外显子缺失或内含子插入。其次,所有结果均可通过RT-PCR进行表征和定量;通过Western印迹鉴定翻译阻断的特性必须等待蛋白质降解,然后才能测定清晰的敲除,但是剪接阻断的mRNA的RT-PCR可以快速检测新的移位带,并且由于RNA与蛋白质相比半衰期短, Morpholino治疗前剪接的mRNA迅速消失。就组成和序列基序而言,最佳的剪接阻断寡核苷酸具有与上述翻译阻断寡核苷酸相同的性质。然而,剪接阻断寡核苷酸具有额外的靶标特异性要求,其中最重要的是具有明确已知的外显子-内含子和内含子-外显子边界的确定的pre-mRNA序列。图6描绘了“典型”的多外显子前mRNA靶标。为了产生外显子缺失,前mRNA中至少需要3个外显子,这样就存在一个内部外显子以及第一个和最后一个外显子。靶向内部外显子的外显子-内含子边界或内含子-外显子边界通常会导致邻接外显子及其侧翼内含子全部或部分缺失,如图7的上部所示。但是,如果存在大量外显子在剪接位点与共有位点的差异上,也可能产生内含子插入。认为发生这些插入是因为在剪接被封闭的外显子侧翼的内含子的远端的未被剪接的边界不能被聚集在一起,这可能是由于小的核核糖核蛋白(snRNPs)的结合不良。靶向前mRNA中第一个或最后一个外显子的剪接点通常会触发完全或部分内含子包涵。我们认为发生这些插入的原因是,当Morpholino寡核苷酸阻止snRNP的正常剪接伴侣结合时,从受阻的外显子接合处跨过内含子的snRNP保持无伴侣状态,并且在第一个末端的远端没有剪接接头或最后一个外显子,其余的snRNP没有可以剪接的另一个剪接点。在图6中,e1i1或i3e4 Morpholinos可能分别引起内含子1或3的插入,i1e2或e2i2 Morpholinos可能引起外显子2的缺失,而i2e3或e3i3 Morpholinos可能引起外显子3的缺失。然而,如上所述,一些期望触发缺失的寡核苷酸可以代替地触发插入,并且如下所述,隐蔽剪接位点的激活可以使插入变成部分插入,或者删除变成部分删除。图6.典型的多外显子pre-mRNA中潜在的剪接连接靶标图7描述了靶向4-exon转录物中exon3-intron3边界的两个最可能的结果。请注意,隐蔽剪接位点可以被激活,并且可以代替图7的第一个示例中所预测的那样剪接至上游剪接位点的5'端。通常,隐蔽剪接位点的激活会导致部分缺失或部分插入。图7.用Morpholino oligo阻断E3I3后观察到的最终产物德雷珀(Draper)BW,莫尔科斯(Morcos),金梅尔(Kimmel)用吗啉代寡核苷酸抑制斑马鱼fgf8 pre-mRNA剪接:一种基因敲低的定量方法。创世记。2001 Jul; 30(3):154-6。图8显示了阻断内部外显子的一系列可能结果,包括外显子跳跃(最常见的结果),内含子插入,以及隐蔽剪接位点激活,部分外显子缺失和部分内含子插入的产物。该图显示了靶向供体部位的吗啉代;尽管由于功能性受体位点的序列受到更大的限制,但不太可能激活隐性位点,但通过阻断受体位点也可能获得相似的结果。图8.通过用Morpholino oligo封闭一个剪接供体而产生的可能产物的例子总之,针对任何剪接连接的Morpholino寡核苷酸预期会产生完全或部分单外显子缺失或完全或部分单内含子插入。要使用RT-PCR分析由阻断剪接的Morpholino产生的mRNA产物,请制作一组针对预期缺失或插入序列外(5'和3')的引物。该基因工具的客服人员会很乐意帮助你俩设计的寡核苷酸,并帮助您确定实验的最佳方法。欢迎致电或发送电子邮件给我们。局限性:使用Morpholino寡核苷酸的最主要限制分为两类,即设计和交付。但是,对设计的限制和交付的挑战只是对无意识的限制。关于设计,只要靶向mRNA或pre-mRNA的适当区域并且避免有问题的部分例如自身互补性和G的游走,则寡核苷酸很可能是有效的。但重要的是要记住,由于吗啉寡核苷酸通过位阻机制起作用,有效的寡核苷酸必须靶向序列以阻断翻译,mRNA修饰前(例如剪接)或调节性蛋白结合(例如内含子剪接抑制剂,外显子剪接)增强剂)。就递送而言,Morpholino主链的不带电性质在历史上一直造成了递送困难,因为Morpholinos无法与阳离子脂质试剂(脂质转染试剂)一起递送。对于培养中的细胞,与使用脂转染试剂来传递核酸相比,使用我们的Endo-Porter试剂传递吗啉提供了良好的传递功效,并且毒性更小。电穿孔也是将吗啉代导入细胞的好方法。当然,已证明将吗啉代蛋白显微注射到胚胎中是非常有效的。应当指出,几乎在所有最广泛研究的生物系统中都已经实现了传递。我们建议您查看我们的出版物数据库,使您能够浏览所有研究的生物。只要您了解良好设计的特征和交付技术并将其计划到实验中,我们相信您将以无与伦比的特异性,最小的非反义作用和惊人的低毒性实现最干净的敲除。